Some time ago, we published this article about how bees use electrostatics to navigate their environment. Today, we will look at another striking example of how electroreception, the capacity to detect electric signals in the environment, can be a powerful adaptation in the animal world. This time, we dive into the world of elasmobranchs, a group of cartilaginous fish that includes sharks and rays. These animals are extremely sensitive to very weak electric fields, which they detect with special organs in their skin, the ampullae of Lorenzini. These organs can detect bioelectric fields, such as the tiny electric signals produced by nearby prey like fish.

{kind=link}
If you would like to learn more about ongoing research and conservation efforts concerning these fascinating animals, you may also want to visit the website of the Dutch Elasmobranch Society (in Dutch: Nederlandse Elasmobranchen Vereniging).
Electrosensitivity in sharks and rays
The first convincing experiments showing that electroreception plays an important role in the sensory perception of elasmobranch fish date back to the 1960s. In a classic study from 1962, Dijkgraaf and Kalmijn described a curious behaviour in spotted catsharks (Scyliorhinus canicula). When the researchers brought a rusty steel wire close to the animals’ heads, the sharks reacted, even when they were blindfolded. From a distance of just a few centimetres, they were able to detect the object and steer away from it. But when the same test was repeated using a glass rod, the outcome was very different. In that case, the sharks only showed an avoidance response once they physically touched the rod. Similar behaviour was later observed in the thornback ray (Raja clavata).
These results led the researchers to suspect that sharks and rays, and likely other elasmobranch fish as well, can detect weak galvanic electric currents generated at the surface of metal objects in seawater. In other words, the animals were not responding to the object itself, but to the tiny electric fields associated with it. Because glass does not conduct electricity, it does not produce such currents, which explains why the fish could not detect it from a distance.
From these experiments, the researchers concluded that elasmobranch fish must possess a highly refined sensitivity to electric fields. In the experiments described in Dijkgraaf and Kalmijn’s paper, swimming sharks and rays showed a strong escape response when exposed to electric fields with a voltage gradient of only about 1 microvolt per centimetre. To put that into perspective, such signals are roughly one million times weaker than the Earth’s atmospheric electric field, discussed in our earlier article on electroreception in bees. Also when compared to the electric fields produced by everyday household electronics, these signals are extremely faint. In fact, they are comparable in strength to the bioelectric fields generated by your own body; for example by the nervous system or the beating heart.
What is sensing electricity?
At first, scientists did not know which sensory mechanism allowed sharks and rays to detect such extraordinarily weak electric signals. The behavioural experiments clearly showed that the animals were responding to electric fields, but the biological “detector” behind this ability remained uncertain.
One extraordinary structure attracted attention: the ampullae of Lorenzini. These ampullary organs form an extensive network embedded in the skin of sharks and rays. Together, the organs constitute an additional sensory system, alongside more familiar senses such as touch, smell, or vision. Each ampulla consists of a bundle of sensory cells grouped in a structure known as a sensory bulb. This bulb connects to the animal’s nervous system and brain. From the bulb, a narrow canal runs outward through the skin and opens to the surrounding seawater via a small pore on the surface. These canals are filled with a jelly-like substance. Interestingly, this jelly has roughly the same electrical resistivity as seawater, but its electrical behaviour resembles that of a semiconductor. This structure is crucial because the canal connects the seawater outside the body to the sensory cells inside. The ampulla can thus detect differences in electric potential, i.e., electric fields, between its external pore and the internal tissues of the shark. Even extremely small voltage differences are enough to trigger a signal to the brain.
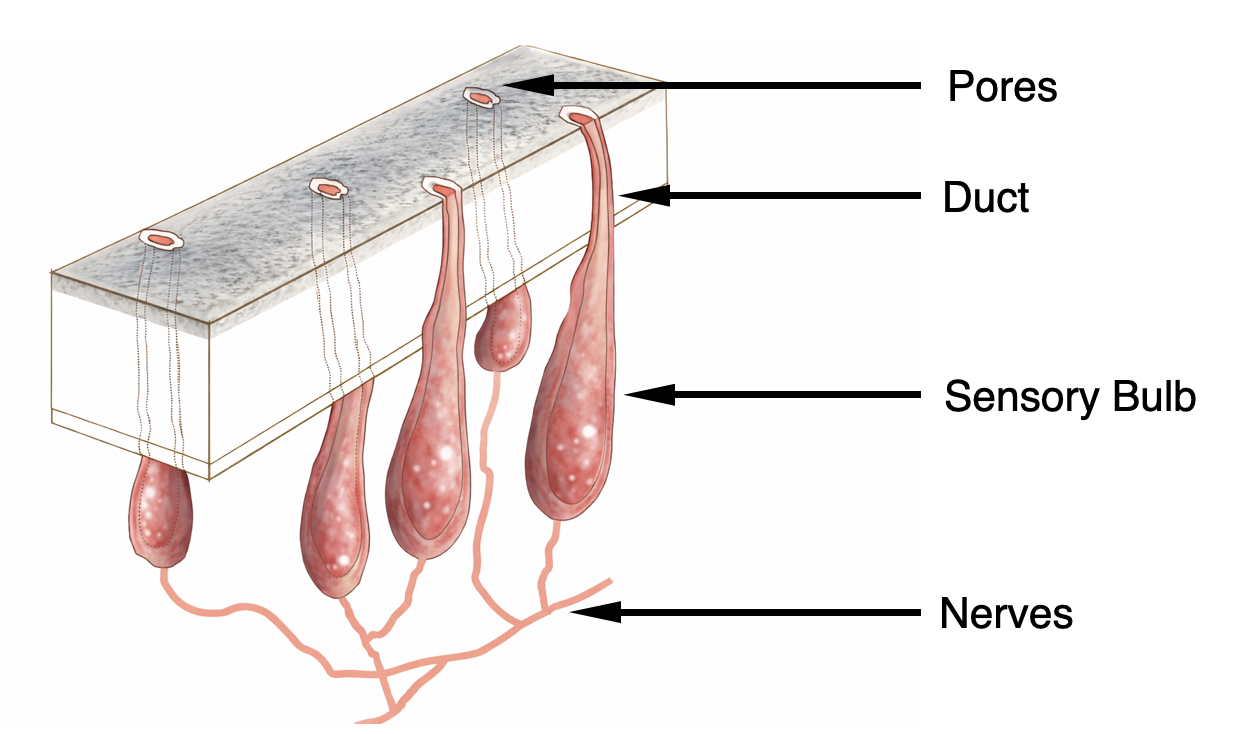
{kind=link}
Although the ampullae of Lorenzini were first described in the 17th century by the Italian ichthyologist and physician Stefano Lorenzini, their true function remained unclear for centuries. The link to electric signals was only firmly established in the mid 20th century, through experiments such as those reported in [Murray, 1960,1962], [Dijkgraaf & Kalmijn, 1962], and later synthesised in [Kalmijn, 1971].
An important clue came from anatomy. In sharks, the ampullary system is concentrated mainly around the head. In the earlier experiments with electrically charged rods, it appeared that only the head region was sensitive to electric stimuli, not the rest of the body [Dijkgraaf & Kalmijn, 1962]. This spatial match between the location of the ampullae and the region of electrical sensitivity strongly suggested that the two were connected.
Even more convincing was what happened when the organs were experimentally damaged. Animals whose ampullae of Lorenzini had been removed or partially destroyed no longer reacted normally to electric fields in their surroundings. From this, researchers concluded that the ampullae are indeed the sensory organs responsible for electrosensitivity in species such as the spotted catshark [Kalmijn, 1971].

{kind=link}
Around the same time, electrophysiological studies (electrophysiology is the branch of the measurement of this flow) provided direct measurements of how these organs respond to stimulation. These experiments showed that the ampullae are sensitive to mechanical and thermal stimuli, but also to extremely weak electric fields. In both catsharks and thornback rays, voltage gradients on the order of typical bioelectric signals, around 1–2 microvolts per centimetre, produced strong neural responses [Murray, 1960, 1962].
With time, the evidence had become overwhelming that sharks and rays are equipped with an electric sense and we now know that the ampullae of Lorenzini function as electroreceptors. [A.J. Kalmijn, 1971]
The life hacks of sharks and rays: How do the Ampullae of Lorentzini help them?
Once it was clear that the ampullae of Lorenzini function as electroreceptors, two obvious questions followed. First, which natural electric fields exist in the habitats of sharks and rays that are interesting to be detected? Second, do the elasmobranchs actually use such fields in an important way, for example when hunting?
The answer to the first question is relatively straightforward. All living animals produce bioelectric signals. These arise from processes such as nerve activity and muscle contractions. In seawater, these tiny electrical signals spread into the surrounding environment. For their experiments, researchers focused on the bioelectric fields produced by the flatfish Pleuronectes platessa (the European plaice, in Dutch: scholvis), a common prey species that shares its habitat with predators such as Scyliorhinus canicula (the small spotted catshark, in Dutch: kleine gevlekte kathaai) and Raja clavata (thornback ray, in Dutch: stekelrog).

{kind=link}
The second question required carefully designed behavioural experiments. In a series of studies [Kalmijn, 1971], elasmobranchs from the North Sea were collected and brought to a laboratory where the animals were kept in shallow plastic pools, large enough for free movement but small enough for close observation. The researchers examined how the small spotted catshark and the thornback ray use electroreception during hunting, not merely for obstacle avoidance or orientation. In the wild, these predators often hunt flatfish that bury themselves in sand to avoid detection. Yet, hungry sharks and rays are remarkably precise hunters, even when their prey is completely concealed.
When a plaice was within about 15 centimetres, a feeding response was triggered, even if the fish was buried under sand. The shark would suddenly but smoothly approach the prey, whirl up the sand covering it, and expel the sand through its gill slits. It would then hold the plaice in its jaws and swim while shaking its head vigorously to tear the prey into swallowable pieces. Rays showed a different, but equally targeted, strategy. After detecting the plaice, a ray would pounce on it and then push the flexible edges of its wing-like pectoral fins against the sandy bottom. This allows for enclosing the prey beneath its body, while the covering sand gets removed by blowing and suction. In this case, the sand is however expelled beneath the ray’s body, since its gill slits are located on its underside. At the same time, the ray made short, jerky turning movements, as if manoeuvring the prey into the right position for capturing it. As an interesting side note, the strongest responses were observed in hungry animals. When they were not hungry, sharks and rays still appeared to register the presence of prey, sometimes approaching briefly or swimming around it, but they often did not bother to attack and were even described as “lazy”.
To isolate the role of electric signals more clearly, the researchers designed an elegant experiment [Kalmijn, 1971]. Living flatfish were placed inside chambers made from agar, a plant-based gelatinous substance that forms a porous gel. The idea was to confine the fish in a way that allowed electric fields to pass through, but minimized other cues such as smell or movement. The agar chambers were then buried under sand in the shark pools.
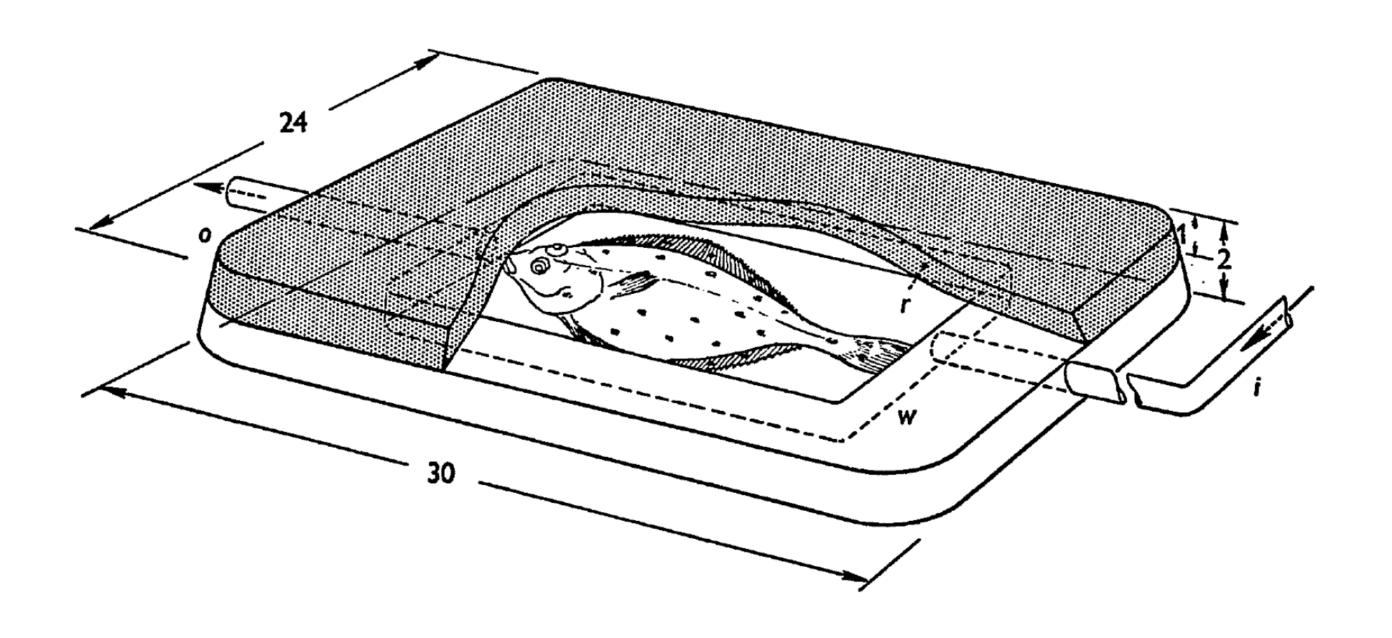
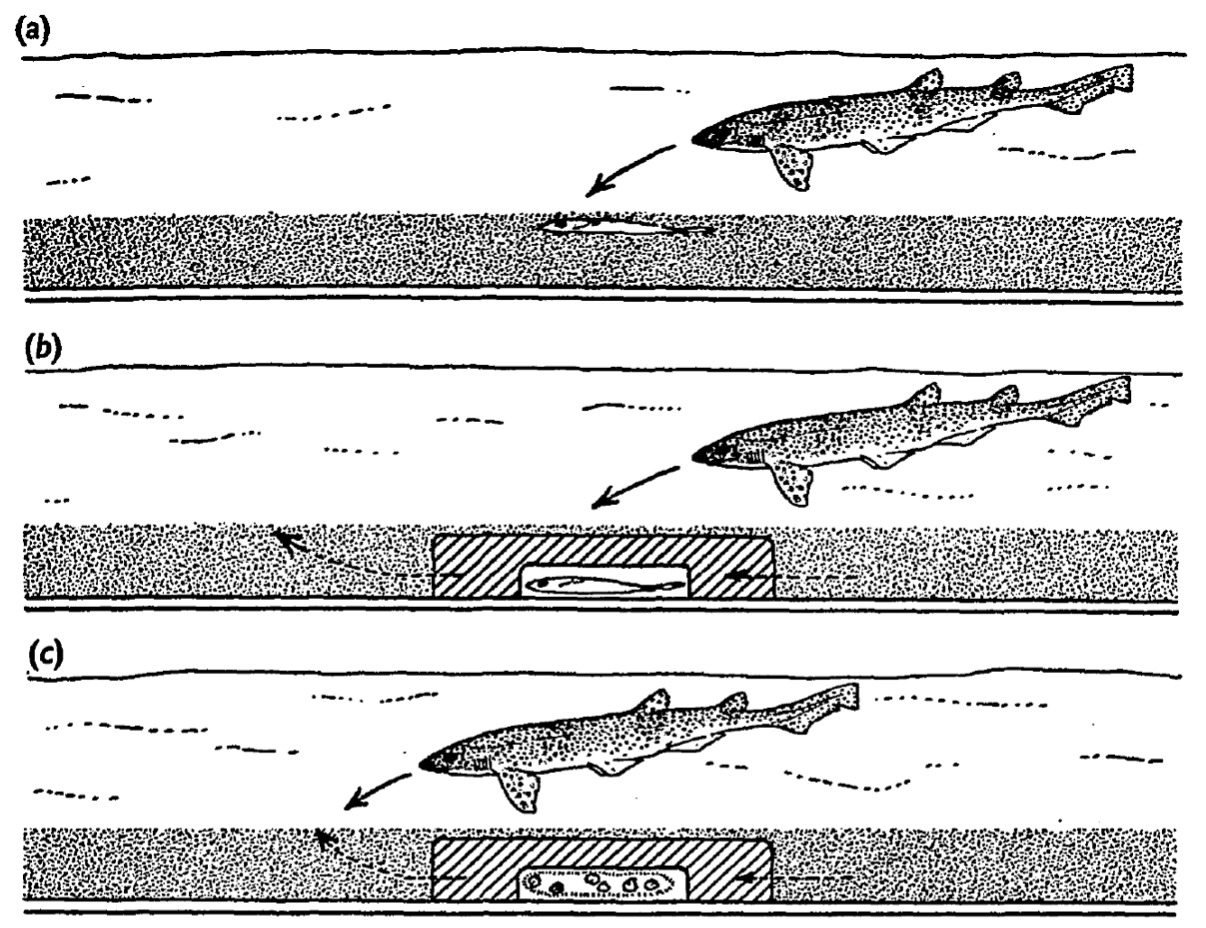
The researchers went even further. They buried small electrodes in the sand to artificially mimic the bioelectric field of a flatfish. Remarkably, sharks and rays responded to these artificial signals in exactly the same way as they did to real living prey. In one particularly revealing test, pieces of fish were placed near, but not attached to, flatfish mimicking electrodes. The sharks’ typically well aimed attacks were directed not at the fish pieces themselves, but at the electrodes. This showed that, at close range, electrical cues can be an even stronger stimulus than the smell of food. In other words, sharks may rely on their electric sense more than their smell, when it comes to the final strike.
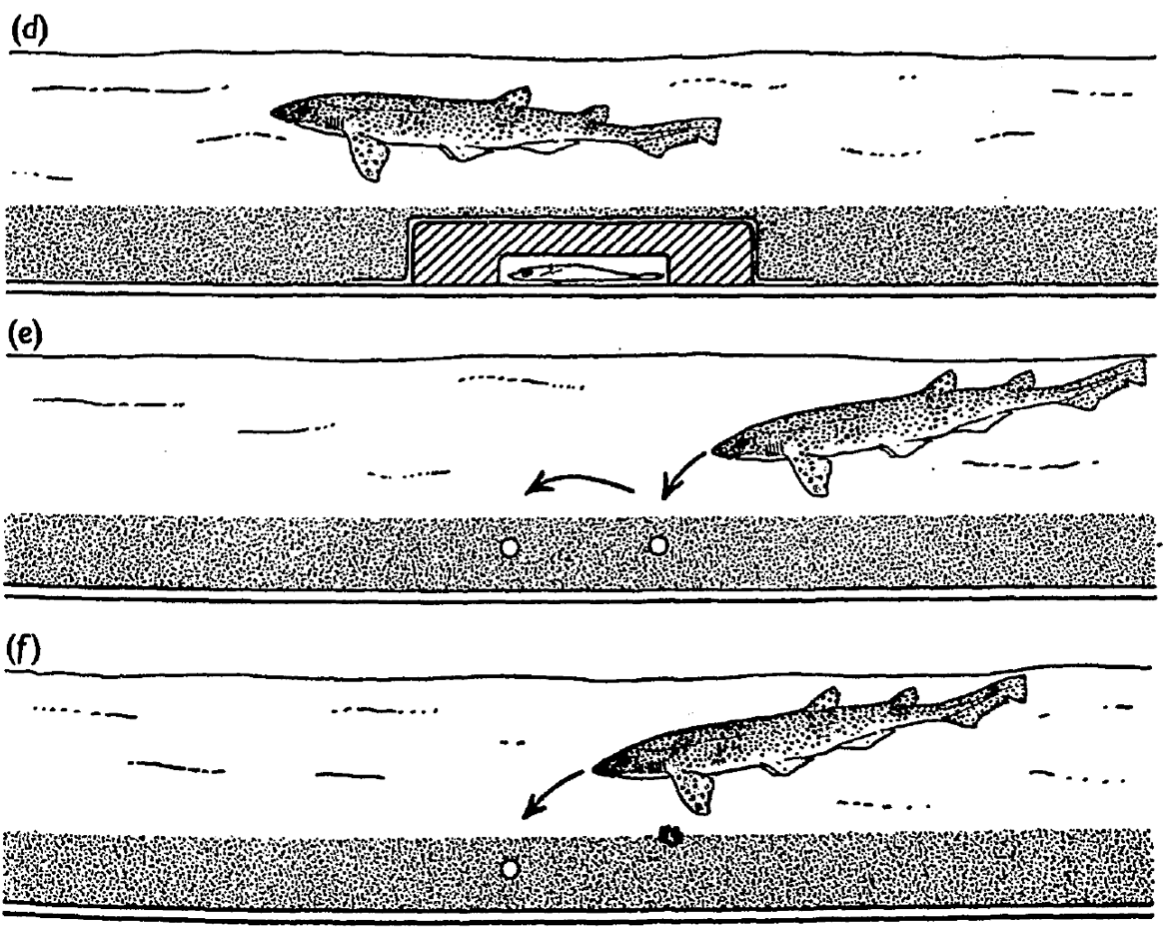
Conclusion
The studies discussed above proved that the ampullae of Lorenzini are specialised organs that detect extremely weak electric signals produced by other animals. This ability is a highly specialised adaptation to hunting, which is central to the evolutionary success of elasmobranchs like sharks and rays.
There is, however, an important limitation. The bioelectric fields generated by animals in seawater decrease very rapidly with distance. Their voltage gradients fall off steeply, which means that electroreception only works over relatively short ranges, typically on the order of about 15 centimetres. Beyond that, the signal simply becomes too weak to be detected.
Within this short range, however, the electric sense is remarkably precise. As the experiments showed, it still functions even when the prey animal is buried in sand or enclosed in an agar container. In such cases where vision is useless and smell may be limited, the electric field remains detectable. [Kalmijn, 1971]
This stands in contrast to odour plumes, for example from a wounded or bleeding fish. Chemical cues can spread over much larger distances and attract predators from far away. At the same time, they are easily disturbed by water currents and rarely allow for a very accurate localisation of the prey. Electroreception and the sense of smell have therefore co-evolved to complement each other. A smell will guide a shark or ray into the general area of the prey, while the electric sense enables a precise final strike.
References:
Murray, R. W. (1960). Electrical sensitivity of the ampullae of Lorenzini. Nature (London), 187, 957.
Dijkgraaf, S., & Kalmijn, A. J. (1962). Verhaltungsversuche zur Funktion der Lorenzinischen Ampullen. Naturwissenschaften, 49, 400.
Murray, R. W. (1962). The response of the ampullae of Lorenzini of elasmobranchs to electrical stimulation. Journal of Experimental Biology, 39, 119–128.
Kalmijn, A. J. (1971). The electric sense of sharks and rays. Journal of Experimental Biology, 55(2), 371–383.